51- Los ojos
Los ojos son órganos sensoriales vitales que nos permiten percibir el mundo visualmente. Son responsables de captar la luz y convertirla en señales eléctricas que el cerebro interpreta como imágenes. Además de su función visual, los ojos desempeñan un papel importante en la comunicación no verbal y expresión de emociones.
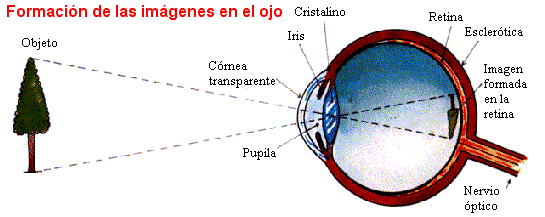
La anatomía del ojo incluye varias estructuras clave:
- Globo ocular: Es la parte principal del ojo y tiene una forma esférica. Está compuesto por varias capas, incluyendo la córnea (la capa transparente en la parte frontal), la esclerótica (la capa blanca y resistente que cubre la mayor parte del globo ocular) y la retina (la capa sensible a la luz en la parte posterior).
- Iris: Es la estructura coloreada en el centro del ojo y está perforada por la pupila, que controla la cantidad de luz que entra en el ojo.
- Cristalino: Es una lente transparente ubicada detrás del iris que enfoca la luz en la retina.
- Retina: Es la capa sensible a la luz que contiene células fotorreceptoras llamadas conos y bastones. Estas células convierten la luz en señales eléctricas que se transmiten al cerebro a través del nervio óptico.
- Nervio óptico: Es el nervio que lleva las señales visuales desde la retina hasta el cerebro, donde se procesan y se interpretan como imágenes.
Además de estas estructuras principales, el ojo también cuenta con músculos oculares que permiten el movimiento del globo ocular y la coordinación de los dos ojos para una visión binocular.
Cuidar la salud de los ojos es fundamental para mantener una buena visión. Esto implica mantener una alimentación equilibrada, proteger los ojos de la exposición excesiva a la luz solar y a pantallas digitales, y realizar revisiones periódicas con un especialista en la salud ocular.
En caso de experimentar problemas visuales o cualquier tipo de molestia en los ojos, es importante buscar atención médica para un diagnóstico y tratamiento adecuados.
51- Los ojos
¿Cómo vemos?. Para ver necesitamos:
- luz (parte de la radiación del espectro solar con longitudes de onda desde el rojo, 750 nm, hasta el violeta, 350 nm),
- que los ojos funcionen bien,
- y que el cerebro interprete bien lo que los ojos le transmiten.
Lo que vemos y cómo lo vemos varía de acuerdo con experiencias personales previas y con nuestra personalidad. Es el toque subjetivo que da el cerebro a la visión.
Más ilusiones ópticas ilusiones óptica

Resulta que las dos son del mismo color, pero las juzgamos diferentes por el contexto en el que aparecen: las verdes, cruzadas por franjas naranjas y las “azules”, por franjas magenta, como explican en Discover Magazine, donde explican que la ilusión está sacada de la web del psicólogo Akiyoshi Kitaoka.
Si pasamos la imagen por Photoshop o por cualquier programa que reconozca el color, podemos comprobar que el código RGB de ambas espirales es 0, 255, 150 (un tono de verde). Pero, como en la ilusión de Adelson, resulta imposible ver las dos espirales del mismo color.

Algunas de las ilusiones ópticas juegan con cómo organizamos nuestra percepción. Cuando la información es ambigua, no sabemos si estamos viendo a un pato o un conejo, o una calavera o una señora, como en esta ilusión creada en 1892 por el artista Charles Allan Gilbert y con muchas versiones posteriores.
Todo está en el cerebro. Las ilusiones ópticas revelan las suposiciones que el cerebro hace sobre lo que en realidad ve.
Identificamos los objetos porque existe una grabación previa en el cerebro. Un ciego que de repente recuperara la vista, no podría identificar ningún objeto.
Veamos a continuación la explicación que da la óptica a la entrada de la información en el cerebro y cómo se determina cuál es la distancia mínima a la que dos puntos deben estar separados para que el ojo los distinga.
Para ver un objeto, este debe reflejar la luz que le llega. No se produce reflexión apreciable de las ondas a no ser que el objeto sea de un tamaño por lo menos de la longitud de onda de la luz que incide. Por lo tanto, con la componente de la luz visible de menor longitud de onda (violeta) podemos ver objetos de un tamaño mínimo de 4·10 -7m.
Se pueden emplear ondas sonoras para «ver» nuestro entorno. El sonar se emplea para la detección de los objetos que nos rodean mediante la reflexión de ondas sonoras. Con sonidos audibles por el hombre el menor tamaño detectable es de 2 cm., pero el murciélago usando ultrasonidos detecta insectos de 2,8 milímetros.
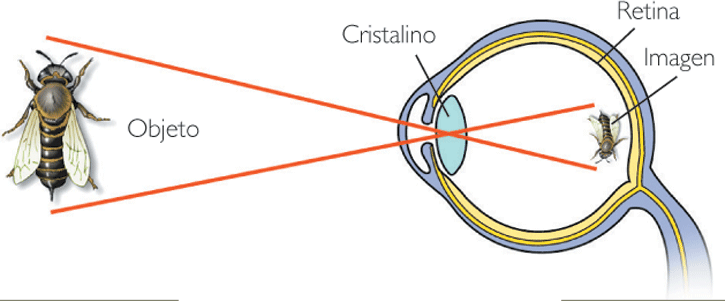
Supongamos que el objeto tiene el tamaño adecuado y se ilumina con luz visible. El sistema óptico del ojo es capaz de recoger los rayos reflejados que divergen del objeto y llegan a él.
El cerebro identifica la posición del objeto como el punto donde convergen las prolongaciones del haz de rayos que le llegan procedentes del objeto.
Para ver nítidamente un objeto la imagen de cada uno de sus puntos debe estar enfocada sobre la retina: los rayos salientes de un punto del objeto que sean captados por el ojo deben concentrarse todos en un mismo punto de la retina, sobre sus células sensibles.
Un objeto se verá borroso si los rayos que parten de él son concentrados por las lentes del ojo delante o detrás de la retina.
Los rayos que llegan reflejados o refractados dan lugar a que pensemos que está en un lugar distinto a donde realmente está, ya que el cerebro interpreta que el objeto se encuentra en la prolongación de los rayos.
Esto da origen al concepto de posición virtual (como si estuviera allí).
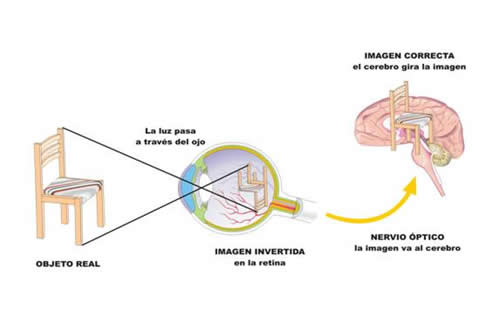
El cerebro debe poner «derecha» la imagen que se forma «invertida» en la retina (arriba es abajo, derecha es izquierda). Utiliza para ello el centro visual situado en el hemisferio izquierdo del cerebro y en su zona posterior.
Cuando miramos un objeto lo vemos siempre bajo un ángulo aparente (a). Este ángulo es el que forman dos rectas que partiendo del iris llegan a los extremos del objeto.
Cuanto más acercamos un objeto mayor es el ángulo aparente con que lo vemos.
Cuanto mayor es este ángulo aparente, mayor es la imagen que se forma en la retina y mayor nos parece el objeto que miramos.
El tamaño de la imagen en la retina (y’) es directamente proporcional a la tangente del ángulo aparente.
El punto hasta el que podemos acercar el objeto sin dejar de ver su imagen nítida y lo más grande posible, es el punto próximo. Es la distancia d, unos 25 cm en el ojo normal. La distancia de la retina al cristalino, D, será de unos 2,5 cm.
Poder de resolución
El ojo tiene un límite para identificar como separados dos puntos próximos. Recuerda que la materia está formada por átomos separados, pero lo que nos muestra el ojo al mirar la materia es un todo continuo.
El poder de resolución se refiere a la capacidad para resolver o distinguir dos objetos que están muy juntos.
Varios factores lo condicionan: el tamaño de las células de la retina, la longitud de onda de la luz y el diámetro de la pupila.
La luz que llega al fondo del ojo atraviesa antes un orificio llamado pupila.
Cuando un haz de luz atraviesa una ranura pequeña se difracta (se abre) y colocando frente a ella una pantalla se observan figuras de interferencia en las que alguna zona queda oscura (luz + luz = oscuridad).
Estudiando la figura de difracción de Fraunhofer producida por una abertura circular sobre una pantalla situada lejos, se obtuvo una relación entre el ángulo subtendido desde el centro de la abertura y el primer mínimo de difracción con la longitud de onda de la luz y con el diámetro de la abertura, Da.
Cuando dos focos puntuales mandan su luz a través de una abertura originan dos diagramas de difracción. Si los puntos están separados, los diagramas se ven separados como una imagen de dos puntos diferenciados, pero si se alcanza la separación angular crítica se ven superpuestos como un único punto deformado.
El ángulo crítico, (q =1,22 l/D, es el formado por los rayos que van del agujero al primer mínimo del diagrama de difracción y al centro del máximo
En ese ángulo está el límite justo para la resolución (criterio de Rayleigh) y pasado este punto la imagen de los dos puntos se superpone.
Cuanto más pequeño sea el ángulo crítico mayor será el poder separador del sistema óptico: más próximos pueden estar los puntos foco y verlos separados. Según puedes observar en la fórmula, esto se puede conseguir disminuyendo la longitud de onda o aumentando el diámetro de la abertura:
La abertura de la pupila es de unos 5 mm de diámetro (Da).
El ángulo crítico para un ojo normal lo dan el diámetro de la pupila y la longitud media de la luz visible (600 nanómetro) y vale:
5 de arco.
Este ángulo mínimo es el ángulo de agudeza visual de esa persona.
Otro factor limitante del poder de resolución del ojo es la separación de los receptores (conos) en la retina debido al tamaño de estas células.
El ojo distingue dos puntos como distintos (separados) cuando la imagen de los mismos se forma en células sensibles distintas. La separación de los conos en la fóvea central es de 1 micrómetro y en otras zonas de 3 a 5 micrómetros que para un globo ocular de 2,5 cm de diámetro da un ángulo de agudeza visual de 2 a 5 minutos de arco.
El Poder Separador del ojo es el valor inverso del ángulo de agudeza visual.
Resumen:
El tamaño de las células condiciona el ángulo de agudeza visual.
Pero la la longitud de onda y el diámetro de la pupila lo determinan también.
Los dos factores conducen al mismo valor. La evolución del sistema de la vista en los humanos, quizás ajustó uno al otro.
Los instrumentos ópticos aumentan el ángulo de agudeza visual y lo que a simple vista parece un punto puede revelarse como dos puntos separados.
Observa que cuando miramos vemos todos los objetos de una escena situados delante, en distintos planos, más o menos nítidos incluso aunque miramos de reojo. Cuando nos fijamos en algo es cuando realizamos un enfoque preciso sobre ese plano. Un paisaje real no es el paisaje plano como el que muestra una fotografía, pero nosotros fundimos todos esos planos en un único plano bien enfocado.
Nuestra visión binocular
Por estar los ojos separados en nuestra cara, cada ojo forma una imagen diferente en la retina. Compruébalo colocando un objeto estrecho centrado y delante de tu cara y cerrando alternativamente los ojos. La superposición que el cerebro realiza de las dos imágenes que los ojos le dan, crea la sensación de relieve.
La siguiente imagen muestra una toma muy exagerada que correspondería a unos ojos mucho más separados de lo normal.
Si la visión que recibe el cerebro por los dos ojos no encaja en sus esquemas lógicos (no la puede componer), se ve forzado a eliminar parte o toda la información que recibe por uno de los ojos para interpretar la escena.
La visión binocular nos permite saber si un objeto es plano o tridimensional. Por el visón binocular podemos distinguir la fotografía de un paisaje del mismo paisaje real. Mirando con un sólo ojo no podríamos hacerlo salvo que nos moviéramos frente a lo que estamos observando.
Cuando miramos una escena tridimensional, aunque lo hagamos con un solo ojo, la posición relativa de los objetos cambia si nos movemos frente a ella.
En la figura inferior desde la posición de la izquierda vemos los puntos A y B con un cierto ángulo y en la de la derecha, al movernos frente a ellos, los vemos alineados.
Si miramos una fotografía desde distintos lugares, la posición relativa de los objetos que en ella están no variará, se mire desde donde se mire.
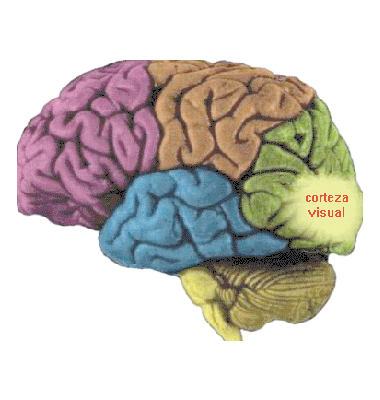
El arribo a la corteza
Pasemos ahora a considerar el sistema visual primario que se proyecta por medio del núcleo geniculado lateral a la corteza visual. Esta vía no contiene otras neuronas, de modo que podemos considerar al núcleo, pequeña parte del tálamo, como una simple estación de relevo. A primera vista, simplemente pasa la información de la retina a la corteza visual en más o menos la misma forma que la recibe. Aclaremos que las neuronas que se proyectan desde la retina son de dos clases: tipo P (por parvo, ‘pequeño’, en latín) y tipo M (por magno, ‘grande’, en latín).oj5Las de tipo P son más numerosas, y diferentes clases de ellas transmiten distintos contrastes entre colores (más bien que los colores mismos, ¡interesante!). Asimismo, diferencias de textura, forma y efectos de profundidad. El tipo M transmiten más rápido y se interesan fundamentalmente por cambios de efectos en el tiempo. Los insumos de los dos tipos de neuronas aterrizan en áreas separadas del núcleo, siendo también discriminados los procedentes de cada uno de los ojos y de la misma manera trasmitidos a la corteza. Podemos comprobar las cuatro líneas que arriban a cada uno de los núcleos en este detalle de la imagen final del artículo precedente.
Pero entremos ya a la ciudadela del cerebro, la corteza. A primera vista, cualquier parte de la corteza cerebral luce como un espantoso enredo, con cien mil neuronas debajo de cada milímetro cuadrado de superficie. Recuérdese que, a pesar de su apariencia corrugada y en forma de bulto, la corteza es en realidad una fina lámina, cuya profundidad se considera su dirección vertical mientras que las otras dimensiones, sus direcciones horizontales, como un pañuelo extendido cuidadosamente sobre una mesa. Recuérdese además la apariencia general de la neurona, semejante a un árbol. Varias capas de neuronas, densamente concentradas en sentido horizontal, resultan comparables a un bosque microscópico de varios pisos. Al visualizarla así, la impresión original de desorden en la corteza desaparece completamente, trocándose en una organización de estratos, como bosques que se repitieran a sí mismos verticalmente.
Seis bosques superpuestos
Después de cuidadosa observación, los neurólogos han llegado a distinguir, para efectos de análisis, seis estratos corticales distintos. En cada uno de ellos ocurren cosas diferentes. El estrato superior (nivel 1) tiene pocos somas, pues consiste principalmente de dendritas apicales de neuronas piramidales de niveles inferiores, junto con los axones de otras neuronas que hacen sinapsis con estas dendritas. Es decir, se trata fundamentalmente de alambrado y de unos pocos cuerpos celulares dispersos aquí y allá. Inmediatamente debajo vienen los estratos 2 y 3, a menudo identificados de manera colectiva como «niveles superiores». Contienen muchas neuronas piramidales. El siguiente estrato, nivel 4, tiene muchas neuronas estrelladas) y unos pocos somas de neuronas piramidales. Este estrato varía mucho en densidad según las áreas corticales, estando completamente ausente en algunas. Los estratos 5 y 6, a menudo identificados colectivamente como «niveles inferiores», tienen de nuevo muchas neuronas piramidales, cuyas dendritas apicales se proyectan hasta el estrato 1.
No solo es el caso que las neuronas varían de estrato en estrato. También, y esto es de gran importancia, las neuronas se conectan de manera diferente. Los niveles superiores (estratos 2 y 3) se comunican solo entre sí, sin proyectarse nunca fuera de la corteza, aunque algunas lo hacen con áreas corticales del otro hemisferio vía el corpus callosum. Algunas de las neuronas del estrato 6 se proyectan de vuelta al tálamo o hacia el claustrum, pequeño satélite de la corteza ubicado inmediatamente debajo de ésta; sin embargo, algunos de sus axones se conectan al estrato 4. La única parte de la corteza cuyas neuronas se proyectan completamente fuera del sistema cortical —esto es, no a otras partes de la corteza ni al tálamo o claustro— son las del estrato 5, con excepción de algunas pocas neuronas conectadas a otras regiones corticales. En ese sentido, puede considerarse que es en este estrato donde la información procesada en la corteza sale hacia otras partes del cerebro y la médula espinal.
Las conexiones que abandonan la corteza, aunque algunas vuelvan a entrar a ella, son siempre excitativas, nunca inhibitorias. Por supuesto, la corteza misma contiene muchas neuronas inhibitorias, aunque mucho menos que excitativas. La razón de estas diferencias estriba en que la inhibición tiene función de control, lo que se necesita mayormente en las computaciones internas de la corteza; en cambio, terminados ya estos cruciales cómputos, lo procedente en adelante es excitar a músculos y glándulas para que hagan su trabajo. La principal, aunque no única, entrada a una región cortical está en la parte superior del estrato 4 o, cuando éste es pequeño o inexistente, en la parte inferior del estrato 3. El estrato 4 conecta principalmente con los niveles superiores, 2 y 3, y éstos a su vez tienen mucha conexión con el estrato 5. Éste tiene conexiones «horizontales» –más bien largas– con el estrato 6 que le subyace, el cual a su vez envía algunos axones «verticales» de nuevo al estrato 4. Hay también importantes insumos de otras regiones corticales para el estrato 1 que hacen contacto con dendritas apicales de las neuronas piramidales de los niveles inferiores.
Si este exiguo resumen de la organización de la corteza parece complicado al amable lector, debemos advertirle que disimula inmensamente su intrincada naturaleza. Puede ser que exista alguna lógica subyacente a tan enorme complejidad, pero puede también ser que su más compacta descripción deba ser la enumeración una por una de sus innumerables conexiones. No lo sabemos todavía. La neocortex, es sin duda la corona gloriosa del organismo humano; pero, reina reservada, no entregará ligeramente sus secretos.
El flujo de la información visual
Como lo vimos en el ensayo anterior, la retina, en el fondo del ojo, comienza ya ella misma el proceso de elaboración del insumo óptico y envía sus resultados a la corteza, a lo largo de dos caminos –el (NGL) y el folículo superior–. Los trozos de información en esta etapa son todavía bastante primitivos, locales y simples. Para que se llegue a producir la percepción tendrán que combinarse y elaborarse mucho más en diversas regiones visuales, localizadas todas ellas en los amplios lóbulos occipitales de ambos hemisferios.
Las células del NGL se proyectan directamente a la (V1), también llamada corteza estriada debido a su apariencia en el microscopio. Si esta región –en seres humanos o monos– se daña en forma importante, el resultado es una ceguera total para la correspondiente mitad del campo óptico –izquierda o derecha–, independientemente de la buena salud de los ojos. Las neuronas del NGL se proyectan a V1 pero, extrañamente reciben igual número de proyecciones de ella que vienen a hacer sinapsis en dendritas un poco alejadas de los respectivos cuerpos celulares. Esta retroalimentación ha provocado especulaciones entre los científicos. ¿Estará relacionada con la conciencia visual? ¿Será la conciencia el resultado de una especie de «hablar consigo mismo» del cerebro? ¡Quién sabe! Agrega a la estupefacción el hecho de que la misma circunstancia se repite en prácticamente todos los circuitos cerebrales: casi todos son circuitos reentrantes, para usar un término común entre los informáticos. Además de estas reentradas reciben también insumos procedentes del tallo cerebral, lo que probablemente significa que el NGL trasmite libremente cuando estamos despiertos, pero su transmisión es bloqueada por el tallo durante el llamado sueño de ondas lentas.
En todo caso, la elaboración de la información visual prosigue con su arribo a V1. Es una región bastante grande y, como excepción a la regla de que hay cien mil neuronas debajo de cada milímetro cuadrado cortical, ese número es en ella cercano a las doscientas cincuenta mil. En toda la región de uno de los hemisferios del macaco se han llegado a contar alrededor de 200 millones de neuronas y aproximadamente un millón de axones que llegan del NGL. Este insumo, todo excitativo, va preferentemente al estrato 4, aunque cierta parte se dirige al 6. El estrato 4 tiene varias subdivisiones. Los ingresos de los estratos P y M del NGL tienden aquí a segregarse también en diferentes sub estratos del nivel 4. Todos los axones entrantes se ramifican extensamente, de modo que cada uno contacta alrededor de mil distintas neuronas. Conversamente, una neurona particular del estrato 4 recibe insumos de muchos distintos axones entrantes. A pesar de esto, solamente una fracción, tal vez 20%, de las sinapsis de una típica neurona estrellada del nivel 4, recibe insumo directo del NGL. El resto de las sinapsis reciben insumo de otras partes, especialmente de axones de otras neuronas de la vecindad. Así, las neuronas del estrato 4 no solo oyen lo que el NGL tiene que decirles, sino que se embarcan en amplios comentarios entre ellas sobre el asunto. Así como el insumo de procedencia retinal se mapea en el NGL, también el insumo de esta procedencia hace lo mismo sobre V1. Este mapa, por supuesto, es de solo un hemicampo visual; además, no es uniforme: como lo esperaríamos, mucho más espacio es dedicado a regiones cercanas al centro de mirada que a la periferia.
La primera región visual se caracteriza porque mucha más cantidad de sus neuronas responden mejor a una pequeña barra de luz o sombra o a un ángulo que a un punto luminoso. La respuesta es también mejor para una línea en movimiento o que se enciende y se apaga. Además, cada neurona tiene una orientación particular de la línea o barra ante la cual más probablemente se dispara. Diferentes neuronas tienen distintas orientaciones preferidas, aunque las inmediatamente encima o debajo tienden a responder a la misma orientación (lo que da lugar a hablar de las «columnas» de la región). Conforme uno se desplaza en la dirección horizontal por la corteza, la orientación preferida tiende a cambiar muy suavemente, aunque pueden darse también discontinuidades abruptas. Sobre un área pequeña de la corteza, digamos de 1 milímetro de diámetro, todas las posibles orientaciones quedan representadas. Muchas otras interesantes especializaciones se dan en las neuronas de esta región, por ejemplo, con relación a centro vs alrededor, o como reacción a la presencia de manchas de un solo color, o al tamaño de una línea (algunas neuronas responden solo si la línea es más bien pequeña o por el contrario si es más bien grande).
Un tipo de neurona muy interesante recibe insumo de los dos ojos y solo se dispara si estos insumos no están exactamente en posiciones correspondientes en las dos retinas. Esto resulta importante para que el cerebro extraiga información sobre la profundidad del objeto en el campo visual, pues objetos a diferentes distancias de la retina producen diferentes disparidades, lo que ofrece fundamento a la visión tridimensional. Algunas neuronas se interesan en una dirección particular del movimiento y no responden al movimiento en la otra dirección. Muchas neuronas responden de la misma manera a cualquier longitud de onda, mientras que otras, que reaccionan ante una mancha, responden mejor al centro que a la periferia de ella, pero su reacción está también condicionada por la longitud de onda. Todas estas neuronas que se especializan en la extracción de alguna característica muy concreta de la imagen se denominan colectivamente «detectoras de rasgos». Entre estos rasgos se cuentan tipos de orientación, disparidades estereoscópicas, longitudes de onda, tamaños de líneas o manchas, etc. Todo lo cual nos sigue convenciendo de lo equivocado que estaban las enseñanzas de los filósofos empiristas del siglo XVIII en el sentido de que podíamos tener impresiones sensoriales «simples».
El Profesor Crick piensa que es razonable proponer, como regla general, que cuando vemos explícitamente algún rasgo, y no simplemente lo inferimos, habrá neuronas en alguna región del cerebro que se disparan estimuladas por él. Considera que si esta regla se confirma habríamos hecho un descubrimiento importante. Por cierto que, desde el punto de vista filosófico, sería una cierta reivindicación del empirismo de Locke y Hume, aunque a un nivel de desarticulación de la experiencia que les sería difícil reconocer como afín a sus propios conceptos. Probablemente semejante a la situación en que se encontraría Demócrito para reconocer los átomos de los elementos químicos actuales como parientes de sus propios pedacitos de materia cuya mezcla consideraba como originaria de todas las cosas.
Para poder comenzar a entender qué parte juega una neurona particular en las operaciones del cerebro necesitamos conocer no solo cuál es su campo receptivo (qué estímulos la excitan) sino también adónde se proyecta, es decir, con cuales otras neuronas hacen contacto las ramificaciones de su axón. Pues ambos aspectos son los que contribuyen a dibujar su contribución estructural, la participación que tienen en la construcción de la mente. En cuanto a las neuronas de la región V1, ellas se proyectan mayormente a la (V2), también un área visual amplia. Como la V1, contiene un mapa de la mitad opuesta del campo visual. Sus neuronas se interesan, en general, por más o menos las mismas propiedades que las de la región V1, tales como orientación, movimiento, disparidad y color, aunque con ciertas diferencias. Casi todas estas neuronas reciben insumos de ambos ojos pero sus campos receptivos son usualmente más grandes que los de V1; además, responden a ellos de maneras más sutiles. Debemos agregar en relación con estas dos regiones que no solo las neuronas de 1 se proyectan a V2; existe aproximadamente un número igual de neuronas de V2 que les pagan la visita proyectándose de vuelta sobre V1. Con una peculiar diferencia: la proyección inicial aterriza sobre todo en el estrato 4 de V2, en tanto que la contra proyección desde V2 evita completamente ese estrato en V1.
Hasta hace poco se consideraban como regiones visuales de la corteza solamente tres, a saber, las áreas de la corteza numeradas de 17 a 19. Las dos primeras, hoy llamadas V1 y V2, las acabamos de describir someramente. Hoy se reconocen adicionalmente otras diecisiete regiones, además de siete que lo son parcialmente. Solo este hecho nos revela la complejidad de los procesos de construcción de la percepción visual. El comportamiento de las neuronas en cada una de esas regiones es diferente, pues cada una tiene distintos insumos y productos. Una de las regiones, V5, es pequeña. Posee un mapa retino tópico bastante bueno del hemicampo visual correspondiente y los rangos receptivos de sus neuronas son típicamente mayores que los encontrados en las regiones V1 o V2. No les interesa el color como tal, pero algunas responden al movimiento de fronteras producidas por contrastes de luminosidad. Esto difiere de lo que pasa en la región V4 que presenta complicadas respuestas a longitudes de onda, pero total desinterés por el movimiento.
Resultados
A continuación, mostramos el diagrama de conexiones neuronales con que culminan las investigaciones del Profesor Crick sobre la localización de la conciencia visual:
Se muestran en la figura las muchas conexiones encontradas entre las diferentes regiones visuales. Se aplica la convención de que cada línea representa muchos axones dirigidos en ambas direcciones, pero con predominio de corrientes en dirección hacia arriba del diagrama. Hemos conservado las iniciales en inglés que designan las diversas estaciones. En particular, al pie de la imagen, RGC (por retinal ganglion cells) representa las células de la retina, LGN es el núcleo geniculado lateral, que se proyecta a la primera región visual, , subdividida en cuatro partes, con la segunda región visual V2 inmediatamente encima subdividida de la misma manera. Los nombres de otras áreas son más o menos arbitrarios y no deben distraer al lector. En la cima, HC representa el hipocampo y ER la sección de corteza (entorhinal cortex) que lleva a él.
En la apreciación de este diagrama sus complicados detalles no deben mortificar al lector. Lo que nos interesa resaltar es la complejidad del proceso neurológico de la percepción visual. El Profesor Crick observa que el diagrama tiene la particularidad de que muy pocas personas habrían podido imaginar que nuestros cerebros estuvieran alambrados de esta manera. Algunas de sus líneas representan millones de axones; otras, tan poquitos como cien mil o menos. Como era de esperar, las áreas contiguas comunican entre sí, con algunas excepciones. Podemos concluir que se da cierta jerarquía, aunque por razones distintas de la contigüidad espacial. Es más bien porque conforme ascendemos, el rango de la recepción neuronal –el tamaño del poquito de mundo exterior a que se refiere cada neurona– aumenta, de modo que en las regiones más altas puede llegar a cubrir todo el hemicampo o incluso ambos hemi campos visuales. Además, conforme más alta la región en el diagrama, más globales y complejos los rasgos que quedan representados.
Lo anterior merece un comentario filosófico. De lo que se trata aquí no es de que conforme avanzamos en la red nos vayamos acercando a la residencia oficial del Mago de Oz, adonde habitaría el homúnculo ante el cual el cerebro haría una presentación teatral que reproduciría –»en subjetivo»– la realidad exterior. Si pensáramos así, no habríamos ganado nada en comprensión ni explicado cómo trabaja el cerebro. Lo que sucede es muy distinto: es el recorrido por los distintos «talleres» de descuartizamiento y posterior ensamblaje de los estímulos iniciales de la sensibilidad lo que va trazando la estructura de la experiencia. Y esta estructura solo obtiene su pleno sentido sobre el trasfondo de las reacciones emotivas y pragmáticas que suscita en otras partes del cerebro y del organismo entero, indispensables para completar la vivencia subjetiva. De ahí la importancia de las conexiones que el Profesor Crick no llegó a explorar y que tendrán que ser exhaustivamente investigadas por otros científicos. La importancia de la obra de Crick es que por fin un científico de talla le perdió el miedo a investigar metódicamente la conciencia y que sus incisivos y rigurosos procedimientos han ofrecido inspiración invaluable a muchos investigadores jóvenes que siguen sus pasos.
El profesor Crick mismo apunta a esa necesidad de ulterior investigación cuando se pregunta por el significado de las conexiones inversas que parecen desandar el camino en toda la trama de la red. Asimismo, sugiere algunas hipótesis que podrían imaginarse al respecto. Podrían ser notificaciones de éxito que tiendan a reforzar las sinapsis usadas para obtener la síntesis superior y hacer así más simples otras síntesis futuras. Podrían tener también que ver con el mecanismo de la atención o con los de la memoria visual, o relacionarse con la sincronización de las oscilaciones neuronales, tema que constituye el foco de atención del siguiente ensayo. O podría ser que sirvieran para que las agregaciones de nivel superior orientaran el proceso en los niveles anteriores. Si esto se confirmara, sería la primera verificación a nivel neuronal del método dialéctico a que nos referimos en otra parte. Podríamos agregar que tal vez fueran avisos de defectuosa transmisión y pedidos de reenvío, como los que imponen los protocolos de Internet. O simplemente ser notificaciones al nivel inferior de que se ha tenido éxito en alguna síntesis y que puede, en consecuencia, ahorrarse ciclos de recomputación en adelante. La práctica de la inteligencia artificial y la experiencia con el funcionamiento de las redes pueden aun sugerir muchas otras posibilidades. En todo caso, la decisión entre todas estas y otras hipótesis solo puede surgir de trabajo experimental y análisis lógico, asignaciones insoslayables para los científicos del futuro.
Todo lo anterior no constituye por ahora más que especulación. Algo queda sin embargo bastante claro para el Profesor Crick y para nosotros:
Existen muchas diferentes regiones visuales, cada una de las cuales analiza el insumo visual de maneras distintas y complejas. No obstante, hasta ahora no se ha podido localizar una región específica en la cual la actividad neuronal correspondiera exactamente al vívido cuadro del mundo que vemos frente a nuestros ojos. Podría pensarse, viendo el diagrama, que ello ocurre en alguna de las más complejas estructuras como el hipocampo y sus estructuras corticales asociadas (marcadas HC y ER), localizadas en la cima de la jerarquía. Pero una persona puede perder todas estas regiones, en ambos lados del cerebro, y todavía reportar que ve bastante bien y de hecho comportarse como si lo hiciese. En resumen, entendemos cómo el cerebro desarma el cuadro, pero no comprendemos suficientemente todavía cómo lo vuelve a armar.
Notas
Nota 1: Las neuronas que se proyectan a los colículos son todas tipo M, lo cual significa que el sistema secundario es ciego al color, lo mismo que las especies que poseen exclusivamente ese sistema visual, como los batracios.
Nota 2: Por este y otros pequeños grandes descubrimientos se otorgó el Premio Nobel a Torsten Wiesel y David Hunter en 1981.
Nota 3: El Profesor Crick acaba de morir, a la edad de 88 años, ocupado todavía activamente en estas investigaciones. Gloriosa muerte para alguien a quien la humanidad debe tanto. Agosto 2004.